
")



da Leadership Medica n. 2 del 2000
Abstract
Recenti progressi immunopatogenetici hanno sottolineato il ruolo del sistema immune come target e protagonista attivo nel corso delle infezioni da Hiv.
In particolare è dato risalto al ruolo della ridotta capacità rigenerativa dei T linfociti, del repertoire del TCR e del possibile ruolo del timo. È anche riferito circa il ruolo importante della terapia HAART nel controllo della viremia, pur evidenziando la necessità di combinare tale terapia con sostanze immunoristoratrici quai le citochine al fine di garantire un ripristino della funzione immune relativamente ai linfociti naive ed alla possibilità di intervenire anche sul reservoir cellulare infetto.
Sommario
- Introduzione
- Attivazione immunologica nelle infezioni da HIV
- Ruolo genetico nella storia naturale dell'infezione
- Timo ed infezione da HIV
- Terapia HAART e ricostituzione immunologica
- Prospettive terapeutiche
- Conclusioni
Introduzione
Negli ultimi anni progressi farmacologici ed immunopatogenetici nonché aspetti epidemiologici hanno trasformato le infezioni da HIV in una malattia cronica, a prevalente trasmissione sessuale e con espressioni patogenetiche caratterizzate da una complessa interazione immunologica in cui coesistono attivazione cronica ed immunodepressione.
In breve, questa infezione è stata trasformata in una malattia dalla maggiore sopravvivenza che interessa non solo categorie a rischio ma, con progressiva incidenza, anche le popolazioni "normali".
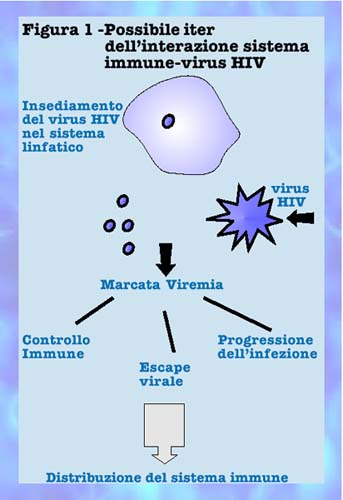
Questa metamorfosi è avvenuta grazie alle migliori e più rapide conoscenze del ruolo del sistema immune e dei suoi complessi networks, che coinvolge lo studio del timo, del repertoire T linfocitario, alla cresciuta importanza dello studio del tessuto linfoide ed alla introduzione delle nuove terapie combinate basate sull'associazione diversa di inibitori delle proteasi, analoghi nucleosidici e non (Figura 1, a sinistra).
Oggi è pertanto opportuno definire l'infezione da HIV come una malattia cronica in cui la storia naturale è determinata dall'equilibrio dinamico tra la capacità di sopravvivenza del virus e la possibilità dell'organismo di neutralizzarlo non escludendo il ruolo di fattori genetici; in questo scenario il sistema immune, pur impegnando tutte le sue numerose risorse, non riesce ad eradicare il virus.
Questi nuovi concetti hanno prodotto una serie di prospettive patogenetiche abbastanza chiare, quali:
a. non esiste una latenza biologica dell'infezione. Il virus, anche se in quantità ridotta, soprattutto nei linfonodi continua a replicare;
b. il sistema immune, sin dalle fasi più precoci dell'infezione, è esposto ad una attivazione cronica;
c. il tipo di risposta immune nelle fasi iniziali dell'infezione (espressione delle molecole HLA, delezione del repertoire T linfocitario, tipo di recettori per le chemochine) può condizionare la storia della malattia;
d. uno stato di depressione cronica di alcune funzioni immuni (linfopenia dei linfociti "naive", linfociti mai venuti a contatto con l'antigene), coesiste con l'attivazione cronica e/o segue, determinando un background assai caratteristico in cui si assiste alla contemporanea presenza di aumento e diminuzione di determinate popolazioni linfocitarie e citochine;
e. il problema della ricostituzione immune dopo la terapia HAART;
f. la possibilità di associare altre terapie a quella antiretrovirale, per una migliore eradicazione del virus dalle cellule "reservoir".
Attivazione immunologica nelle infezioni da HIV
L'infezione primaria da HIV determina una massiccia replicazione virale che diffonde il virus agli organi linfoidi, caratterizzando una risposta immune sia umorale sia cellulare molto vigorosa, che, tuttavia, non impedisce allo stesso virus di continuare a replicare in tessuti inaccessibili al sistema immune come il SNC, i macrofagi e le cellule "resting", ossia non in fase attiva ove il virus è integrato come provirus DNA. Tanto caratterizza, soprattutto, la fase asintomatica dell'infezione.
Si ritiene, in base a modelli matematici, che ogni giorno siano prodotti almeno 109 virioni e più del 99% di tale produzione proviene da cellule neo-infette, la cui emivita è di circa 1-6 giorni.
La viremia presente, dopo circa 6 mesi dall'infezione, può essere un criterio predittivo di progressione, ed è influenzata sia da fattori correlati al sistema immune (citochine, chemochine e loro recettori, molecole HLA) sia al virus (capacità replicativa, tasso di mutazioni, citopaticità).
In queste fasi iniziali, il sistema immune fenotipicamente esprime i segni della attivazione mediante:
- presenza di numerosi T-linfociti CD8 citotossici specifici per il virus ed attivati;
- espressione di markers di attivazione specifica dei T linfociti (recettore CD69; recettore per la IL-2; recettori per la transferrina);
- produzione abnorme di citochine Th1, ossia pro-infiammatorie, progressivamente sostituite da citochine Th2, anti-infiammatorie, che possono essere dannose ai fini di una induzione di efficacia della immunità cellulare.
A tal riguardo, recentemente, Autran et al. hanno definito il ruolo di alcuni linfociti, dotati del recettore CD7+ , i linfociti CD4+CD7+, che si associano ad un profilo secretorio citochinico di tipo Th0/Th1, mentre i linfociti CD4 non dotati di tale recettore, i linfociti CD4+CD7-, sarebbero associati ad un profilo prevalente di tipo Th2.
Su queste popolazioni, anche il nostro gruppo di Immunologia (Perrella et al.) da tempo lavora ed ha prodotto una serie di dati preliminari relativi al loro ruolo nelle infezioni HIV (dati non pubblicati).
Questa equazione, attivazione immunologica=maggiore progressione, è infatti osservata nei soggetti Africani ove le coinfezioni da HIV con malattie parassitarie endemiche determinano una più rapida evoluzione dell'infezione.
L'attivazione del sistema immune non è capace di eradicare l'infezione ed a lungo termine risulta dannosa causando un collasso della funzione immune.
Da tali premesse, si evince che la distruzione dei linfociti CD4+ più che effetto di un danno patogenetico sembra epifenomeno di una potente attivazione immunologica e dei suoi numerosi mediatori tra cui i linfociti CTL (in analogia ad altre importanti patologie umane come l'epatite cronica B) e diversi mediatori quali le citochine.
Ruolo genetico nella storia naturale dell'infezione
E' noto che il genere umano è caratterizzato da un'elevata variabilità individuale e che la maggiore o minore capacità di resistere alle infezioni è condizionata da differenti assetti genici ed, in particolare, degli aplotipi HLA.
L'analisi degli eventi immuno-virologici associati all'infezione primaria ha suggerito che una differente espansione delle regioni Vbeta sui linfociti CD8+ causa una differente espressione della risposta citotossica HIV-specifica più o meno favorevole sull'evoluzione della malattia.
La scoperta recente che alcuni geni conferivano una quasi totale immunità alle infezioni da HIV all'1% di soggetti di razza caucasica e che alti livelli di chemochine avevano un potere soppressivo sulla replicazione del virus ha rafforzato il concetto del condizionamento genetico sull'espressione e prognosi della infezione.
Queste scoperte hanno fornito nuovi elementi alla costruzione di un più corretto modello patogenetico che prevede la presenza di co-recettori (per le chemochine) per la penetrazione del virus nella cellula target. Questi co-recettori sono utilizzati, normalmente, dalle chemochine. Notevoli dubbi esistono sul fatto che alti livelli di chemochine siano protettivi per l'infezione da HIV, mentre non esistono dubbi che uno stato di omozigosi (raro) per il deficit del gene CCR5 conferisce una protezione nei confronti del virus HIV.
Timo ed infezione da HIV
La funzione del timo declina progressivamente con l'età come confermano studi bioptici e radiologici. Nei soggetti HIVAb+, la residua funzione del timo è rapidamente annullata come è osservato anche nei bambini con tale infezione.
L'economia del timo nell'elaborazione della risposta immune è essenziale perché:
- genera i linfociti naive e CD8+;
- assicura un'ampia eterogeneità del repertoire dei recettori per l'antigene (TCR).
In caso di continua distruzione dei linfociti naive senza una nuova generazione, per il mantenimento dell'omeostasi linfocitaria, l'organismo può provvedere con l'espansione periferica di cellule già esistenti.
In breve, per mantenere costante il livello di linfociti, si assiste ad una divisione di quelli esistenti; ma questo comporta una diminuzione dei livelli di eterogeneità del repertoire dei T-linfociti, ossia della funzione di riconoscere molti antigeni.
Nei soggetti HIVAb+, la funzione timica può essere controllata mediante lo studio dei livelli di TREC (rearrangement excision circles) che identificano le cellule recentemente emigrate dal timo tra le cellule T circolanti.
Nel corso della vita, questi livelli diminuiscono proporzionalmente con l'età. In pazienti con infezioni da HIV, i livelli di TREC sono risultati più bassi rispetto a quelli descritti in controlli sani di pari età.
Tanto, oltre a suggerire che la funzione timica continua anche nell'età adulta, fornisce un contributo importante al possibile ruolo di marcatore dei TRECs.
Ciò é suggerito anche da studi di McCune et al, ove il trattamento con farmaci antiretrovirali ha indotto un aumento di TREC anche se tale linea di evidenza non sembra, comunque, confermata da altri studi.
Queste ipotesi confermano che alla distruzione dei linfociti CD4+, l'organismo risponde con diversi meccanismi nel tentativo di mantenere costante il pool di linfociti circolanti. Sino a poco tempo fa, la più seguita ipotesi circa la patogenesi della linfopenia CD4+ sosteneva che il livello dei linfociti CD4+ era il risultato di un equilibrio tra distruzione e produzione e che, a lungo termine, le capacità riproduttive si esaurivano determinando la grave deplezione dei linfociti CD4+.
Questa ipotesi è stata contraddetta da una serie di evidenze sperimentali (Hellerstein et al.) che hanno sostanzialmente suggerito che il declino dei linfociti CD4+ era causato da una più breve sopravvivenza e da ridotta capacità riproduttiva. Infatti in prove sperimentali che utilizzarono una tecnica di marcatura non-radioattiva fu dimostrato anche da Pantaleo et al. che:
- la quantità di linfociti CD4+ distrutti non correlava alla nuova produzione di linfociti;
- la produzione di cellule CD4+ nei soggetti HIV Ab+ non è significativamente diversa, nelle fasi precoci, da quella dei soggetti HIV-negativi, essendo il turnover appena 2-3 volte più elevato.
Alla luce di questi recenti studi, l'equazione distruzione=ricostituzione non sembra più accettabile emergendo che la limitata capacità di rinnovare le cellule è uno dei meccanismi possibili della progressiva distruzione del sistema immune.
Terapia HAART e ricostituzione immunologica
Molti studi hanno evidenziato che la terapia combinata antiretrovirale esercita un ruolo importante nella patogenesi delle infezioni da HIV, migliorando l'omeostasi linfocitaria e riducendo la replicazione virale.
In particolare, lo studio pilota di Autran dimostrò che la terapia HAART produceva un precoce aumento delle cellule memoria (CD4+CD45RO+) ed un molto tardivo incremento dei linfociti naive (CD4+CD45RA+) oltre al declino dei linfociti attivati CD8+, ossia i CD8+CD38+, nei soggetti in fase avanzata di malattia.
Questi dati non sono, in seguito, stati condivisi da altri autori, ed anche il nostro gruppo di ricerca dell'Ospedale D.Cotugno (Perrella, Guarnaccia, Atripaldi, D'Antonio, Canonico e De Luca) in cooperazione con il gruppo di Immunologia dell'Università Federico II (Racioppi, Matarese) ha evidenziato con alcuni dati preliminari presentati in occasione di "International Conference on Emergence of Infectious Diseases ", Napoli 1998, che l'incremento dei linfociti naive non era significativo dopo 12 mesi di terapia, mentre l'incremento dei linfociti memoria era confermato sin dalla 4a settimana di trattamento.
Il nostro studio (attualmente submitted in AIDS) (Tab.I) era effettuato sia sul sangue periferico sia su sospensioni cellulari ottenute da biopsia linfonodale ed era relativo all'analisi delle cellule memoria, naive, dei linfociti CD8+CD38+ e di un nuovo immunofenotipo, i linfociti CD4+CD7+ , che è correlato ad un profilo citochinico di tipo, prevalentemente, Th1.
Questo studio evidenziava:
- significativa riduzione dei linfociti CD8+CD38+ nel sangue periferico rispetto ai linfonodi;
- i linfociti naive non erano diversi nei due distretti;
- i linfociti CD4+CD7+ erano diminuiti nei due distretti.
In sintesi da questi dati preliminari è possibile ricavare che:
- l'aumento, almeno nei primi mesi, di terapia è correlabile ad una ridistribuzione dei T linfociti dal tessuto linfoide alla periferia;
- l'attivazione T linfocitaria è solo diminuita in periferia ma persiste a livello dei linfonodi;
- nei nostri pazienti, dopo 12 mesi di terapia, non era evidenziata alcuna ripresa della popolazione naive.
Questa considerazione era suffragata anche da un altro studio del nostro gruppo, ove l'analisi del repertoire T linfocitario evidenziava una migliore eterogeneità nei linfonodi rispetto al sangue periferico;in altri termini, questa ridistribuzione garantirebbe una migliore qualità funzionale dei T linfociti versus un maggior numero di antigeni, determinando la riduzione dell'incidenza di patologie infettive opportunistiche.
Un altro aspetto importante della terapia HAART è legato alla sua durata: è noto che una sospensione può rapidamente determinare un rebound della viremia.
Infatti, secondo diversi studi, anche piccole quantità di provirus DNA integrato nel genoma cellulare, possono indurre una massiva replicazione virale.
Un limite ulteriore di questa terapia è determinato, inoltre, dalla sua inefficacia nei confronti del reservoir cellulare infetto, ossia di quelle cellule che sono in fase di quiescenza e sfuggono all'azione dei farmaci ed infine dall'emergenza di ceppi resistenti.
Questi limiti lasciano spazio a possibili terapie alternative quali:
1. manipolazioni genetiche sul gene del recettore CCR5;
2. utilizzazione di citochine nella ristorazione del sistema immune.
Prospettive terapeutiche
La durata ed il costo dei farmaci antiretrovirali rappresentano dei problemi irrisolti ad alto impatto sia in termini di prognosi sia di Sanità Pubblica e ciò costituisce uno dei compiti a cui è chiamata a rispondere l'autorità regionale sanitaria che si snoda attraverso l'interazione tra l'Assessore alla Sanità, on.Ettore Liguori, l'Area di Assistenza Sanitaria, coordinata dal Dr. Roberto Pepe ed il Cerifarc, scientificamente coordinato dallo scrivente.
Uno dei prossimi obiettivi è la formulazione, da parte del nostro gruppo, di un protocollo di terapia combinata tra farmaci antiretrovirali, ovvero di associazione tra IL-2 e la terapia HAART .
Gli obiettivi dello studio sono essenzialmente rivolti a:
1. valutazione dell'associazione in termini di ristorazione immunologica;
2. impatto sui livelli del reservoir di cellule naive infette;
3. migliorare la prognosi a lungo termine;
4. ridurre la durata totale della terapia HAART.
La metodologia diagnostica sarà effettuata mediante la determinazione monitorata delle seguenti variabili:
a. analisi degli immunofenotipi T linfocitari circolanti (naive, memory, CD4+ ,CD8+);
b. valutazione dello stato di attivazione dei linfociti CD8+;
c. analisi del repertorio T linfocitario mediante la metodica di "Spectratyping" basata sulla determinazione del TCR e delle sue regioni variabili;
d. misurazione dei livelli plasmatici di citochine Th1 e Th2 e della Leptina;
e. livelli di TRECs mediante PCR Real Time;
f. analisi quantitativa dell'HIV RNA circolante mediante RT-PCR.
Conclusioni
Nonostante i numerosi progressi, la immunopatogenesi dell'AIDS ed una terapia definitiva restano ancora da definire.Sembra chiaro che il sistema immune non svolge solo un ruolo passivo di target dell'infezione ma anche un ruolo attivo espresso dalla stimolazione sull'amplificazione virale tramite la mediazione di citochine e chemochine.
Nello scenario di una contemporanea presenza di attivazione e depressione del sistema immune non può essere escluso il concorso di fattori genetici ed il ruolo dei fattori virali espressi da quasi-species assai eterogenei.
Oreste Perrella
Primario Infettivologo
Azienda Ospedaliera "D. Cotugno"
Direttore Scientifico del Centro Riferimento AIDS della Regione Campania
Bibliografia
1. Fauci AS. Nature 384, 529, 1996
2. Fauci AS. Science 262, 1011, 1993
3. Perrella O. in: Infezioni HIV: aggiornamenti e prospettive. Ed. Cesil 1995
4. Perelson AS et al. Science 271, 1582, 1996
5. Mellors JM et al. Science 272, 1167, 1996
6. Wei X et al. Nature 373, 117, 1995
7. Coffin JM. Science 267, 483, 1995
8. Poli G et al. J Exp Med 172, 151, 1990
9. Clerici M et al. Immunol Today 14, 107, 1993
10. Autran B et al. J Immunol 154, 1408, 1996
11. Kovacs JA et al. N Engl J Med 335, 1350, 1996
12. McCune JM et al. J Clin Invest 101, 2301, 1998
13. Ameisen JC et al. Immunol Today 12, 102, 1991
14. Clerici M et al. Immunol Today 15, 575, 1994
15. Margolick JB et al. Nat Med 1, 674, 1995
16. Pantaleo G et al. Proc Natl Acad Sci USA, 94, 254, 1997
17. Connor RI et al. J Exp Med 185, 621, 1997
18. Lusso P et al. J Virol 69, 3712, 1995
19. Premack BA et al. Nat Med 2, 1174, 1996
20. Paxton WA et al. Nat Med 2, 412, 1996
21. Pantaleo G et al. Nat Med, 5, 27, 1999
22. Hellerstein M et al. Nat Med 5, 83, 1999
23. Autran B et al. Science 277, 112, 1997
24. Li TS et al. Lancet 351, 1682, 1998
25. Fleury S et al. Nat Med 4, 794, 1998
26. Perrella O et al. AIDS 8, 395, 1994
27. Perrella O. et al. J Infect Dis 168, 1079, 1993
28. Bhatia M et al. Nat Med 4, 1038, 199